TÉMATA:
Obecná stavba stěny trávicí trubice. Elektrická a mechanická aktivita hladké svalové buňky v GIT. Obecné typy kontrakcí hladké svaloviny GIT. Obecné typy pohybů GIT. Složení a funkce slin. Složení, funkce a řízení sekrece žaludeční šťávy. Složení a funkce pankreatické šťávy. Enterický nervový systém – anatomie.

Obecná stavba stěny trávicí trubice
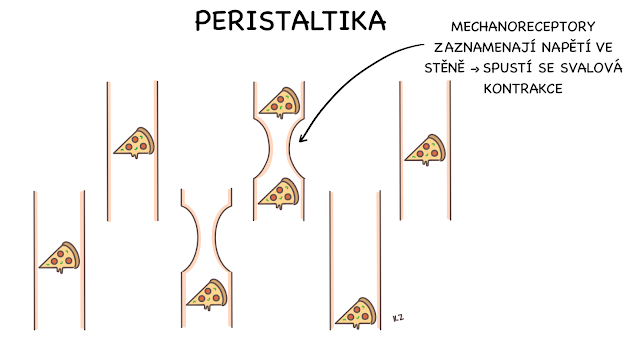
Stěna trávicí trubice má ve finále jednoduchou stavbu, z čehož ale vybočuje pár výjimek (např. žaludek). Skládá se celkem ze čtyř vrstev. Nejvnitřněji najdeme sliznici (tunica mucosa), na kterou nasedá podslizniční vazivo (tela submucosa). Dále je zde svalovina (tunica muscularis), která má vnitřní vrstvu cirkulární (stratum circulare) a vnější vrstvu longitudinální (stratum longitudinale).
Když dojde ke stažení cirkulární svaloviny, střevo se zúží. Naopak při kontrakci longitudinální svaloviny se střevo zkrátí a rozšíří. Na tomto principu spolupráce svalových vláken funguje peristaltika. Poslední vrstvou je vrstva povrchová – adventitia nebo serosa – podle toho, v jakém úseku trávicí trubice se nachází. Serosa je součást viscerálního peritonea, a pokrývá tedy části nacházející se intraperitoneálně. Adventitia je pouze řídké vazivo, které funguje jako spojení s okolními orgány mimo peritoneum (najdeme ji např. na jícnu).

Ve stavbě trávicí trubice najdeme hned dvě vegetativní nervové pleteně enterického nervového systému (viz dále). V podslizničním vazivu se nachází plexus submucosus Meissneri a mezi vrstvami svaloviny leží plexus myentericus Auerbachi.
Enterický nervový systém

Sympatikus, parasympatikus a také enterický nervový systém (ENS) jsou součásti autonomního nervového systému. ENS funguje v podstatě samostatně, proto ho také někdy nazýváme jako druhý mozek nebo mozek střev. Jeho hlavní úlohou je kontrola funkce trávicího systému, což je také důvod, proč se nachází přímo mezi vrstvami trávicí trubice.
Plexus submucosus Meissneri, nacházející se v podslizničním vazivu, řídí sekreci látek do trávicí trubice a také její krevní zásobení. Trávicí trubici ovlivňuje spíše na místní úrovni (na rozdíl od plexus Auerbachi).
Plexus myentericus Auerbachi je pleteň, kterou nalézáme mezi cirkulární a podélnou vrstvou svaloviny. Je zodpovědná zejména za peristaltiku střev a ovlivňuje rozsáhlejší část trávicí trubice.

Info navíc: V trávicím traktu nalézáme také tzv. Cajalovy intersticiální buňky, ty můžeme popsat jako “střevní pacemaker”, který udává rytmus střevní peristaltiky.
Elektrická a mechanická aktivita hladké svalové buňky v GIT
Leiomyocyty (hladkosvalové buňky) GITu musí být sehrané pro správné fungování, a proto tvoří funkční soubuní, což znamená, že se tento shluk buněk chová tak, jako by byl jednou buňkou, i když pochopitelně o jednu buňku nejde. Toto chování jim umožňuje zejména vzájemné propojení pomocí nexu – “gap junctions”, díky kterým si buňky mohou předávat elektrický signál v podobě iontů. Motilita GIT je řízena nejen enterickým nervovým systém, který je schopen fungovat i bez vnějšího nervového zásobení (in vitro), ale i in situ přeci jen spolupracuje s ostatními částmi ANS – parasympatikem a sympatikem, které zde fungují trošku jinak, než jsme zvyklí z jiných částí těla.

Zkráceně… Parasympatikus v GIT téměř vše stimuluje a sympatikus naopak inhibuje. Pamatujete si na popis sympatiku “fight or flight” a popis parasympatiku “rest and digest”?
Parasympatikus tedy podporuje motilitu střev, abychom mohli správně trávit. Když ale bude trávenina procházet střevem, zavazely by ji cirkulární svěrače, které se tam nachází. Proto jsou parasympatikem inhibovány. Sympatikus funguje opačně. Tak, jako téměř v celém těle, i tady způsobuje sympatikus vazokonstrikci. Dostatečné prokrvení je však důležité pro správnou tvorbu trávicích enzymů a správnou resorpci. Opět tedy vidíme, že sympatikus trávení inhibuje.

Motilitu GIT můžeme rozdělit na dva základní typy dějů – pohyby a kontrakce. Pohyby jsou výsledek kontrakce.Tyto děje jsou zodpovědné za promíchávání, posun a skladování tráveniny v gastrointestinálním traktu. Kontrakce dělíme na tonické, které udržují určité napětí (např. v žaludku nebo tračníku) a rytmické, při kterých pozorujeme pravidelné střídání kontrakcí a relaxací. Pohyby pak dělíme na propulzivní, kde je hlavním zástupcem pohyb peristaltický, který též označujeme jako myenterický reflex, neboť je pro jeho uskutečnění nutná koordinace pomocí myenterické nervové pleteně. Dalším typem pohybu je pohyb mísící, což je pohyb jednotlivých orgánů k promísení tráveniny s trávicími šťávami.
A co ta elektrická aktivita?
Leiomyocyty v GITu mají speciální elektrické vlastnosti, které nazýváme BER (nebo nech být; haha, no nic… Jdeme dál!). BER je zkratka pro bazální elektrický rytmus, tzv. pomalé vlny. Tento rytmus generují pacemakerové buňky (Cajalovy), na který pak nasedají tzv. hrotové potenciály (něco jako akční potenciál, ale s delším trváním a menší amplitudou). Důležité je ale vědět, že žaludeční svalovina se dokáže stáhnout i bez hrotového potenciálu – stačí jí pouze malá vlna.
Pro správnou činnost trávicího systému je potřeba, aby byly do lumen vylučovány sekrety, které zastávají funkci zvlhčení potravy (usnadnění polknutí, obalení hlenem), což usnadňuje hladký průchod trávicí trubicí a zároveň ji chrání před poškozením. Dále zastávají funkci trávicí (díky enzymům), imunitní funkcí nebo mají také vliv na řeč (ve smyslu artikulace).
Takové sekrety jsou v GIT tvořeny buď žlázovými buňkami ve sliznici nebo specializovanými žlázami, kam patří slinné žlázy, žaludeční žlázy, exokrinní pankreas, žlázky jícnu a střeva a také játra.
Předtím, než začneme diskutovat o tom, co a jak v jednotlivých částech funguje a co se vyplavuje, bychom si měli uvědomit 2 věci…
- Všechny části GITu něco secernují.
- Všechny sekrety mají společný základ (voda, ionty z plazmy, HCO3 a hlen).
Složení a funkce slin
Slina (saliva) je produktem slinných žláz dutiny ústní. Jedná se o tři velké žlázy – glandula parotis, submandibularis a sublingualis. Dále máme nespočet malých žlázek, které jsou rozesety po dutině ústní. Úkol slin není životně důležitý, ale jejich přítomnost výrazně ovlivňuje hygienu a komfort dutiny ústní a jazyka, navíc se významně podílí na artikulaci.
V ideálním případě se vytvoří 1 – 2 litry slin za 24 hodin. Toto číslo je však orientační a jeho hodnota může být ovlivněna hydratací organismu, přítomnosti různých vůní a chutí, přežvykováním, nevolností (pozitivní vlivy), strachem, únavou nebo stavem spánku (negativní vlivy).
Nyní je vhodné zmínit, jak je vlastně produkce slin regulována. Jde hlavně o regulaci nervovou – konkrétně o ANS. Žlázy, které produkují sliny, mají receptory pro acetylcholin (muskarinové receptory (M) – parasympatikus, α a β adrenergní receptory – noradrenalin – sympatikus; dále substance P či VIP (kotransmiter acetylcholinu)). Nezapomeňme, že účinek parasympatiku je výrazně silnější. Po navázání transmiterů na receptory, je další aktivita v rukou druhých poslů cAMP a IP3.
Parasympatikus působí pomocí IP3, čímž zvýší influx Ca2+ iontů. Pomocí Ca2+ dependentní kinázy dochází ke zvýšení vodivosti luminálních Cl- kanálů, což způsobí eflux iontů Cl- do lumen a tvorbu negativního potenciálu. Po elektrickém gradientu se do lumen dostává Na+ a H2O, což je důležité pro tvorbu řídkých a vodnatých slin s lehce zvýšeným obsahem α-amylázy (za to může právě vzestup Ca2 + iontů, které se podílejí také na exocytóze vezikulů s amylázou). Parasympatikus působí také vazodilatačně – zvyšuje tvorbu kallikreinu, který štěpí kininogen. Ten se nachází v plazmě a je spojen s bradykininem – působí lokální vazodilataci, čímž zvyšuje průtok krve žlázou.

Sympatikus má vliv přechodný a přes β receptory působí na tvorbu cAMP, přičemž dochází k fosforylaci proteinů a aktivaci enzymů -> tvoří se hustá mucinózní slina s vyšším obsahem α-amylázy a mucinu.
Konečně se dostáváme ke složení slin. Sliny jsou složeny majoritně z vody, elektrolytů a organických látek. Mezi elektrolyty řadíme Cl-, Ca2+, K+, Na+, HCO3-. Organickými látkami jsou mucin, α-amylázy, jazyková lipáza, lysozym, imunoglobulin A a laktoferin. Sekundární slina je hypotonická a její pH se pohybuje v rozmezí 7–8 (v klidu je spíše kyselejší).
Proč tomu tak je? Slina je tvořena ve dvou stupních. Nejprve se vytvoří primární slina ultrafiltrací krve. Je tvořena acinárními buňkami a její iontové složení je víceméně podobné složení plazmy. Při následném přesouvání primární sliny odvodnými kanálky dochází k úpravě jejího složení pomocí buněk vývodů a stává se z ní slina sekundární. Ta je následně vyloučena do dutiny ústní.
Primární slina se tvoří v epiteliálních buňkách acinu, kde je základním dějem transcelulární transport Cl- iontů. Pojďme si představit buňku, která je za tento děj zodpovědná. Na svém basolaterálním povrchu má Na/ K ATPázu, která vytváří hnací sílu pro další transportér, respektive se jedná o sekundární aktivní kotransport Cl-, K+, Na+ do buňky. Dále nacházíme také K+ kanál. Pojďme na luminální stranu. Jak jsme si řekli, Cl- se vylučuje do lumen, kde se tvoří negativní luminální potenciál. Co jsem však ještě nezmínil je to, jak je to vůbec možné. Dochází k tomu tak, že se transport uskutečňuje pomocí kotransportu s HCO3- ionty – oba po koncentračním spádu, což přidává negativnímu luminálnímu potenciálu na negativitě. Po tomto elektrickém gradientu se dostávají ionty Na+ z intersticia do lumen, paracelulárně přes tight junctions. A nebyla by to voda, kdyby po svém osmotickém gradientu nenásledovala Na+. Kromě výše zmíněného Cl- a HCO3- kotransportéru se na luminální straně nachází kanál pro K+.

Primární slinu bychom měli, ale její složení neodpovídá složení sliny sekundární (definitivní)… Bohužel. Je to kvůli tomu, aby mohla být slina dopravena na cílové místo jménem ústní dutina. K tomu potřebuje projít odvodnými cestami, které mají směrem do svého lumen orientované epitelové buňky. Ty zajišťují resorpci iontů a mění tak složení a osmolalitu slin. Podstatou této resorpce je vytáhnout maximální množství
Na+ a Cl- iontů z primárních slin a nahradit je K+ a HCO3- ionty. Jak se to ale děje? Na povrchu lumen máme kanály pro Na a Cl, výměníky Na+/H+, Cl-/HCO3, H+/K+ (první iont směřuje do buňky a druhý do lumen).
Na basolaterálním povrchu nalezneme zase Na+/K+ ATP-ázu, která se podílí na tvorbě hnací síly, protože víme, že se vypuzují 3 ionty Na+ směrem ven a 2 ionty K+ směrem do buňky. Z toho vyplývá, že resorpce Na+ na luminální straně bude vždy větší než sekrece K+. To vytváří v lumen negativní potenciál, který způsobí, že se Cl- vyměňuje za HCO3-. Kromě Na+/K+ ATP-ázy nacházíme na basolaterálním povrchu také K+ a Cl- kanály a H+/Na+ výměník (H+ putuje ven z buňky).

Abychom se však také dotkli otázky pH slin a jejich hypotonicity… Jde o to, že se v odvodní části nacházejí těsné tight junctions, kterými se do lumen nedostává Na+ ani voda. Sekrece HCO3- a K+ iontů do lumen zvyšuje pH slin.
Rychlost sekrece slin můžeme uvádět v různých jednotkách, jako základní rychlost se však uvádí 1 ml/ min/g žlázy. Tato rychlost může vzrůstat, což ovlivní i činnost žlázy. Složení sekundárních slin tak bude více připomínat složení slin primárních.

Funkce slin je provázána se samotným složením. Kromě vlivu na žvýkání a polykání potravy slouží sliny na oplachování chuťových receptorů, čištění zubů a podílejí se také na utěsnění dutiny ústní u kojenců při kojení. Zahajují trávení škrobů (ptyalin a alfa-amyláza – ta působí v pH od 4 do 11, jelikož je v potrava v ústní dutině relativně krátkou dobu, její hlavní účinnost nastává v žaludku, dokud její pH neklesne pod 4; zajímavým zjištěním je fakt, že amyláza dokáže rozštěpit až 50% škrobu, zbytek dokončí až pankreatická amyláza). Také zajišťují imunitní ochranu nebo udržování pH dutiny ústní pomocí HCO3-.
Poznámka: Výše jsme zmínili, že se před zvracením zvyšuje produkce slin. Je to proto, že se na ochraně dutiny ústní před kyselou žaludeční šťávou podílí právě složka HCO3-, proto před zvracením pozorujeme zvýšenou produkci slin.
Složení, funkce a řízení sekrece žaludeční šťávy
V žaludku se nacházejí tubulární žlázky, což jsou v podstatě vchlípeniny žaludeční sliznice bez vývodů, které secernují složky žaludeční šťávy do lumen žaludku. Denně se takové šťávy vytvoří přibližně 2,5 litru.

Složení žaludeční šťávy je podobné všem sekretům GITu. Máme tam vodu, soli, HCl, pepsin, vnitřní faktor a mucin.
Samotné tubulární žlázy dělíme do tří skupin podle toho, kde se nacházejí. Jde o oblast subkardialní (produkce mucinu), oblast fundu (produkce HCl a vnitřního faktoru) a oblast pylorickou (produkce mucinu a gastrinu).
Poznámka: Na začátku a na konci žaludku se produkuje mucin. Je tomu tak proto, aby byly proximální a distální části GIT chráněny před kyselým prostředím, které je v žaludku nezbytné pro trávení.
Je potřeba si uvědomit, že podkladem sekrece jsou buňky, které jsou pro žaludeční žlázky výstelkou. Každou učebnicí je tato část vykládána trošku odlišně a my jsme se tedy rozhodli pro vysvětlení této části využít informace z Borona. Ten si žaludek rozdělil na corpus a antrum. U kardie je zmíněno, že je zbavena parietálních buněk, což souvisí s tím, že se v této části bude tvořit hlavně mucin.
Corpus je tvořen hlavně parietálními a hlavními buňkami. Parietální buňky vylučují HCl a vnitřní faktor (důležitý pro příjem vit. B12 v ileu). Hlavní buňky zase produkují lipázy a pepsinogen, který je pomocí nízkého pH aktivován na účinný pepsin (endopeptidáza = hydrolyzuje vnitřní peptidické vazby). Žaludeční lipáza je důležitá zejména pro kojence při trávení lipidů z mléka. Kromě nich se ve žlázách nacházejí také mucinózní buňky a endokrinní buňky (ECL – histamin).

Antrum neobsahuje parietální buňky. Proto se zde nemůže tvořit HCl ani vnitřní faktor. Místo toho se zde nacházejí buňky hlavní (pepsinogen) a endokrinní (G – gastrin, D – somatostatin)
Kromě těchto speciálních buněk se v žaludku nachází také povrchový epitel, který secernuje HCO3- a mucin.
Poznámka: Mucin chrání sliznici žaludku před chemickým a mechanickým poškozením. Je tvořen z 95% vodou, dále fosfolipidy, proteiny, Na+ a K+ ionty. Mezi vrstvu mucinu a epiteliální povrchovou vrstvu jsou vylučovány HCO3- ionty z epiteliálních buněk, které pufrují HCl.
Významnou funkci v žaludku zastává právě HCl, která zajišťuje snížení pH, díky čemuž se neaktivní pepsinogen může proměnit na účinný pepsin. Dochází také ke znemožnění inaktivace některých vitamínů nebo zajištění baktericidního účinku právě díky nízkému pH. Dochází k také konverzi nerozpustného CaCO3 na CaCl2 nebo redukci Fe3+ na Fe2+, který se lépe vstřebává, napomáhá resorpci proteinů a taky kolagenu – tím, že ho depolymeruje.
Jak HCl v žaludku vzniká?
Než se na toto téma vrhneme, pojďme si představit buňku, ve které se tvoří (tedy parietální buňku). Ta má trojúhelníkový tvar, centrálně uložené jádro, mitochondrie a velmi zajímavou apikální část, která se po stimulaci dokáže morfologicky měnit. Ve stavu klidu tvoří membrána tubulovezikulární systém, kde se v cytoplazmě nachází zásoba membrán s obsahem H+/K+ ATPázy. Ty jsou schopny se začlenit do apikální membrány, díky čemuž se zvětší její povrch a charakter se mění na kanalikulární s vysokým počtem H+/K+ ATPáz (H+ jde z buňky ven a K+ dovnitř). Pojďme se ale už pobavit o samotné tvorbě HCl, která se skládá ze dvou atomů. A přesně tak na ní budeme nahlížet.
Sekrece H+ do lumen začíná na známé ATP-áze – Na+/K+ ATP-áze, která vnáší do buňky 2 atomy draslíku. Draslík se tedy do lumen dostane přes kanál na luminální straně, kde je pomocí již zmíněné H+/K+ ATP-ázy vložen do buňky (výměnou za H+). Ale odkud ten H+ vlastně je? I na to je docela jednoduchá odpověď. Pochází z H2CO3, která se vytváří pomocí karboanhydrázy z CO2 a H2O. Z této reakce zůstane v buňce HCO3-. A co Cl? Ten se do buňky dostane výměníkem HCO3- (z buňky) a Cl- (do buňky). Z buňky je pak vylučován do lumen díky elektrochemickému gradientu.
Platí tedy, že za jeden H+, který bude vyloučen do lumen, dostaneme jeden Cl- iont. Kromě tohoto antiportu HCO3- / Cl- se do buňky může dostat také symportem s Na+. Jednalo by se o sekundární aktivní transport, který je uskutečňován po gradientu Na+, který je odčerpáván z buňky díky Na+/K+ ATP-áze.
Poznámka: V textu jsme zmínili to, že se Cl- do lumen dostane po svém elektrochemickém gradientu. Právě elektrickou část tohoto gradientu má na starosti K+, proto se někdy můžeme setkat s vyobrazením vchodu K + a Cl- do lumen jako se symportem.
Jak se tvorba HCl reguluje?
Nejprve se podívejme na to, co dokáže sekreci HCl v parietální buňce nastartovat. Na parietálních buňkách nacházíme více typů receptorů, které reagují na podněty z okolí. Rozhodně nesmíme opomenout H2 receptory. Ty reagují na histamin, který je produkován žírnými buňkami v žaludku, jež někdy nazýváme ECL buňkami. Histamin působí přes druhého posla cAMP. Dále je to acetylcholin. Ten reaguje s muskarinovými M3 receptory a funguje přes druhého posla IP3. Nejmenší význam má gastrin, ale nemůžeme jej opomenout. Všechny tyto dráhy nakonec končí u různých proteinkináz, které fosforylují H+/K+ výměník. Díky tomu začne pracovat rychleji.
Je třeba zmínit také působky, které sekreci HCl dokáží inhibovat. Takovým působkem je somatostatin nebo prostaglandin E (PGE) Pro fajnšmekry… Děje se to tak, že oba působky účinkují přes Gi (G inhibiční protein), který inhibuje adenylát cyklázu. Takže žádné cAMP a žádná fosforylace výměníku.
Při vyvolávání sekrece žaludeční šťávy jídlem rozlišujeme 3 fáze regulace – podle toho, kde k vyvolání sekrece dochází. Zmiňujeme fázi cefalickou (zrak, čich, chuť), gastrickou (distenze žaludku, peptidy a AMK) a intestinální (distenze duodena, peptidy a AMK).
- Cefalická fáze

Začínáme cefalickou fází, k jejíž stimulaci může docházet nepodmíněně (zrak, čich, chuť) nebo podmíněně (naučeně). Tato fáze je zajišťována X. hlavovým nervem (nervus vagus), což má za následek 4 fyziologické jevy. Zaprvé bude n. vagus působit přímo pomocí acetylcholinu na parietální buňky, přes M3 receptory. Zadruhé bude acetylcholin působit na ECT buňky. Ty následně vypustí histamin, který vyvolává odpověď na H2 receptorech parietálních buněk. Zatřetí se cestou n. vagus, po jeho propojení na postgangliových neuronech, bude v antrum uvolňovat GRP (gastrin releasing peptide). Ten bude podle názvu stimulovat G buňky, které vypustí histamin, aby následně působil na parietální buňky – přímo i nepřímo (díky stimulaci ECL buněk). Ty podporují tvorbu histaminu, který bude opět zaregistrován H2 receptory na parietálních buňkách. Čtvrtou reakcí vagu je jeho vliv na inhibici tvorby somatostatinu v endokrinních buňkách typu D.
Zajímavost: I když má vagus tak pozitivní vliv na sekreci HCl, u některých závažných případů žaludečních vředů se může přistoupit k vagotomii. Ta se provádí velmi selektivně, jinak by mohl zákrok vést k dalším nežádoucím účinkům – zpomalenému vyprazdňování žaludku nebo průjmu. Kromě toho by vás mohl zajímat také příspěvek Ivana Petroviče Pavlova ke zkoumání cefalické fáze stimulace trávení. Pavlovič navíc objasnil fakt, že k vyvolání zvýšení sekrece žaludeční šťávy cestou vagu přispívá také hypoglykémie (navozená inzulinem), a za svůj výzkum na psech dostal v roce 1904 Nobelovu cenu za fyziologii a medicínu.

2. Gastrická fáze
Nastává v době kdy je potrava přivedena do žaludku. Ten se začne plnit a roztahovat se, což má za následek dvě věci. Zaprvé bude aktivován vagovagální reflex a zadruhé dojde také k aktivaci lokální odezvy ENS. Vagovagální reflex bude působit stejnou odezvou, jak jsme si popsali v cefalické fázi (ty 4 efekty). Na úrovni ENS bude odezva obdobná. Bude docházet k uvolnění acetylcholinu a GRP. Další efekt, který do gastrické fáze patří, je efekt peptidů nebo AMK (samotné proteiny by efekt neměly). Jde o to, že v průběhu trávení se proteiny natráví a vznikají peptidy a AMK, které podněcují sekreci gastrinu G buňkami. Ten pak podněcuje vyloučení HCl parietálními buňkami.

Zajímavost: Pokud se nad tím zamyslíme, je to regulace pozitivní zpětnou vazbou – nízké pH v žaludku denaturuje proteiny. Vznikají peptidy, které podněcují sekreci gastrinu G buňkami. Gastrin vyvolává sekreci HCl v parietálních buňkách a cyklus začíná znovu. Na tomto principu tedy funguje i to, proč například kofein, pivo nebo víno (nehledejte za tím složitou chemii, jednoduše snížíme pH – pivo a víno mají pH kolem 3–4 a káva kolem 4–5 🙂 ) dokáží podporovat trávení.
3. Intestinální fáze

Jako poslední tu máme intestinální fázi. Ta nastává ve chvíli, kdy bude potrava ze žaludku přecházet do duodena. Tato fáze je trošku složitější a není podrobně prozkoumána. Co ale víme, je to, že distenze duodena pracuje stejně, jako jsme již popisovali ve fázi gastrické. Podobnost můžeme hledat také v tom, že v této fázi peptidy interagují s G buňkami. Je to součást enterogastrického reflexu, který kromě toho, že zvýší mechanickou aktivitu antra, bude ještě částečně působit na stimulaci sekrece. Nezapomeňme zmínit, že v této fázi bude již převažovat inhibiční efekt, který je zprostředkován řadou faktorů – snížené pH, přítomnost MK, hypertonie chymu (tráveniny) v jejunu nebo ileu. To se děje pomocí lokálních enterogastrických hormonů nebo parakrinních působků.
Složení a funkce pankreatické šťávy

Slinivka břišní je morfologický velmi podobná slinných žlázám, ale oproti nim má navíc ještě přídavnou endokrinní funkci. Vývody slinivky se spojují do jednoho hlavního vývodu – ductus pancreaticus major, který vyúsťuje na Vaterské papile (papilla duodeni major), díky které může být pankreatická šťáva uvolňována do lumen duodena.

Jaké je tedy složení pankreatické šťávy? Obsah pankreatické šťávy se dá rozdělit do dvou fází, a to fáze vodní a fáze enzymové. Vodní fáze je složena z vody a iontů, ze kterých je nejdůležitější HCO3- iont. Enzymová fáze je tvořena enzymy ve formě zymogenů (proenzymy), které se aktivují až ve střevě. Kromě nich se ve šťávě nacházejí i jiné působky (inhibitory trypsinu, prokolipázy, …). Tento sekret je, na rozdíl od slin, izotonický s krevní plazmou.
Exokrinní částí se produkuje pankreatická šťáva, které se vytvoří asi 1 litr. Zde se, jako u slinných žláz, setkáváme s buňkami acinárními a buňkami vývodů. Můžeme tedy říci, že se sekrece pankreatické šťávy také skládá ze dvou fází. Jak k tomu tedy dochází?
První fáze probíhá v acinárních buňkách. Acinární buňky vylučují nejen ionty, ale také enzymy. Enzymy jsou vylučovány formou proenzymu a činností buňky se vytvářejí a skladují zymogenní granula, která jsou vylučována do lumen. Co se týče iontů, hlavním aktem je vyloučení Cl-, které vytváří negativní potenciál v lumen, po kterém (přes tight junctions) do buňky prostupují ionty Na+ a voda po elektrochemickém gradientu. Tato sekrece je zodpovědná za cca 25% objemu pankreatické šťávy.

Ve druhé fázi probíhá výměna iontů Cl- za HCO3- ionty. Mechanismus, který je za toto zodpovědný, je dost složitý. I tak by bylo dobré mrknou na tzv. Cl- recycling. Je to dráha, kterou si buňka vytvořila pro nezávislost výměny Cl- na těch Cl- iontech, které jsou uvolňovány acinárními buňkami. Funguje to tak, že buňka má na svém apikálním povrchu aniontové kanály, které umožňují cirkulaci Cl- iontů mezi lumen a buňkou. Důležité je neopomenout to, že nejdůležitějším souborem těchto kanálů je právě známý CFTR kanál (Cystic Fibrosis transmembrane conductance channel), který je u cystické fibrózy defektní a vede tak k poruchám sekreční činnosti slinivky břišní (snížená pufrovací schopnost, což vede ke snížení pH chymu a následné nefunkčnosti enzymů). Nemění se zde však koncentrace Na+ a K+ iontů (na rozdíl od slin), čímž se zachovává izotonicita s krevní plazmou. Právě 75% objemu pankreatické šťávy je tvořeno ve vývodech natolik, jak je tight junction pro vodu a ionty v této oblasti prostupný.

Na místě je zmínit také to, jak se bude měnit složení šťávy v závislosti na rychlosti produkce. Nebojte se – je velmi jednoduché si to představit. Čím více se nám bude tvořit šťávy, tím více bude zpomalená výměna Cl- za HCO3-. To však není na škodu. Pokud k tomu dochází fyziologicky, tedy tak, že bude větší množství chymu, způsobí nám to také přenos většího množství HCl ze žaludku do duodena, které se tak musí pufrovat. Právě díky zpomalené výměně Cl- a HCO3- bude mít pankreatická šťáva pufrovací schopnost vyšší.
Jak je to ale vše regulováno?
Sekrece pankreatické šťávy je řízena cestou humorální a nervovou. Za nervovou regulaci zodpovídá vagus. Jde o to, že k pankreatu přináší sympatickou (tlumivou) a parasympatickou (stimulační) inervaci. Acetylcholin z parasympatiku odpovídá za sekrece zymogenních granulí. Humorální regulace je zajištěna převážně dvěma hormony a to sekretinem a CCK. Oba působky jsou tvořeny sliznicí duodena. Sekretin se tvoří v S-buňkách díky reakci na snížení pH, následně se vypustí do krevního řečiště, kudy doputuje do slinivky. Tam potencuje sekreci pankreatické šťávy v buňkách vývodů (potencuje tedy zejména sekreci HCO3- a obohacování šťávy o vodu).
Cholecystokinin (CCK) je aktivován jídlem – hlavně lipidy, respektive mastnými kyselinami. CCK se majoritně podílí na sekreci zymogenních granul v acinárních buňkách (a to jak přímou, tak nepřímou cestou). Přímá cesta je pomocí CCKa receptorů, které zvyšují intracelulární hladinu Ca. Tedy zvyšují sekreci zymogenních granul pomocí Ca dependentní proteinkinázy. Nepřímo je to uskutečněno na základě aktivace parasympatických vláken. Proto můžeme tvrdit, že CCK je důležité zejména pro sekreci enzymatické pankreatické šťávy.

Teď už víme, co, kde a jak působí. Ale co to stimuluje? Sekreci pankreatické šťávy lze rozdělit na 3 fáze aktivované jídlem. Je to cefalická fáze, gastrická fáze a fáze střevní.
Cefalická fáze je totožná s cefalickou fází žaludku. Při podnětech dochází k reflexní stimulaci vagu, který uvolní acetylcholin. Acetylcholin působí na acinární buňky vývodů. Tato fáze je zodpovědná za 20% produkce pankreatické šťávy.
Gastrická fáze je zajištěna zejména napnutím mechanoreceptorů v žaludku. Ty vyvolají vagovagální reflex – produkci acetylcholinu ve slinivce a vyloučení gastrinu v žaludku. Ten však nehraje významnou roli pro sekreci pankreatické šťávy.
Důležité je také zmínit, že gastrická fáze nemá signifikantní vliv na sekreci pankreatické šťávy. Poslední (a zároveň nejvýznamnější) fází je fáze střevní. V té se uplatňuje fáze sekretinu vlivem CCK. Je to díky snížení pH, ke kterému dochází po přechodu chymu ze žaludku do duodena. To aktivuje tvorbu sekretinu v S buňkách, což vede k obohacování pankreatické šťávy o ionty HCO3- a vodu. Přitom dochází také k tomu, že v duodenu reagují MK a peptidy s buňkami, což vede k sekreci CCK, který způsobí sekreci zymogenních granulí.
Dosud jsme se v článku zabývali sekrecí enzymů v neaktivní formě. To ale není vše! Více se dozvíte v následující části.
Zajímavost: Ve více studiích bylo zjištěno, že lidské acinární buňky nedisponují receptory pro CCK, což tedy znamená, že aktivace acinárních buněk CCK je majoritně uskutečňována nepřímou cestou.
Na závěr se ještě podíváme na funkci pankreatické šťávy. Už jsme se bavili o tom, že pomocí HCO3- iontů dochází k pufrování pH v duodenu po tom, co do něj vnikne chymus ze žaludku. Neděje se to však jen pro zmíněnou potravu, ale také proto, že enzymy pankreatu mají své funkční optimum v rozmezí pH 7–8. Právě správná produkce HCO3- tedy zajišťuje správné rozložení potravy a následnou absorpci. K tomu, abychom mohli potravu rozložit, jsou potřeba pankreatické enzymy, které jsou nepostradatelné pro správné trávení nebo vstřebávání živin. Do enzymové složky pankreatické šťávy řadíme více druhů enzymů. Jedná se o proteolytické enzymy (trypsin, chymotrypsin, karboxypeptidáza A a B nebo elastázy), lipolytické enzymy (lipáza, fosfolipáza, cholesterolesteráza), amylolytické enzymy (alfa amyláza) nebo nukleolytické enzymy (ribonukleázy a deoxyribonukleázy).
Zajímavou skupinou jsou proteolytické enzymy zejména proto, že jsou secernovány v neaktivní formě. Je to z toho důvodu, aby po cestě nezničily vývodné cesty pankreatu. Abychom je mohli aktivovat, použijeme střevní enterokinázu, kterou produkují enterocyty. Střevní enterokináza aktivuje trypsin a ten následně další proenzymy (chymotrypsinogen, prokarboxypeptidáza A a B, proelastáza). Doposud jsme se vždy bavili o zymogenech a proenzymech, ale jak jsme si už také stihli říct, ne všechny enzymy jsou uvolňovány v neaktivní formě. Ano, mám na mysli enzymy k štěpení sacharidů a lipidů.
Zajímavost: Pankreas uvolňuje ještě jeden enzym, který je určitou pojistkou na ochranu při předčasné aktivaci trypsinu mimo dvanáctník. Jde o tzv. trypsin inhibitor, který dokáže inhibovat aktivitu trypsinu.



